
本文Modulating cortical excitability and cortical arousal by pupil self-regulation發表于2025年5月的 Nature Communications(IF=15.7),來自蘇黎世聯邦理工學院的 Marieke Lieve Weijs 等人為本文的第一作者。
研究背景
大腦的覺醒狀態(即中樞覺醒)影響各種正在進行的高級認知過程,并受多個神經調節核控制,包括去甲腎上腺素能(NA)、藍斑核(LC)以及多巴胺能和膽堿能等。通過利用藍斑(中央覺醒的關鍵調節器)和瞳孔動態之間的機制聯系,已經證明了受試者可以通過基于瞳孔的生物反饋方法獲得對包括藍斑在內的覺醒調節中心的意志控制。本研究將基于瞳孔的生物反饋與單脈沖經顱磁刺激、腦電圖和聽覺oddball任務相結合,探究基于瞳孔的生物反饋是否調節皮質興奮性、皮質覺醒和P300反應的電生理標記。
研究方法
01 實驗一
實驗1探究基于瞳孔的生物反饋結合單脈沖經顱磁刺激(TMS)研究瞳孔自我調節如何影響運動誘發電位(MEPs)測量的皮質興奮性(圖1)。
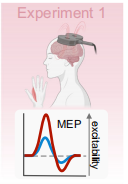
圖1 基于瞳孔的生物反饋技術與單脈沖經顱磁刺激(TMS)相結合,旨在探究瞳孔自我調節如何影響通過運動誘發電位(MEP)測量皮質興奮性。
所有受試者都經歷了2種調節(上調和下調)和1個靜息控制條件:(i)上調,即放大瞳孔;(ii)下調,即縮小瞳孔;(iii)靜息控制條件,受試者的任務是保持自己思想的自由漫游,不使用調節策略。每個條件重復兩次(即總共6個block),條件的順序是偽隨機的,并在受試者之間進行平衡。每組由10個試次組成,從1秒的指示階段開始,給出自我調節或對照條件的提示,然后是4秒的基線測量。然后,受試者應用訓練過的心理策略來調節瞳孔大小15秒,或者休息。在自我調節或靜息階段,輸出兩個經顱磁刺激脈沖到初級運動皮層,第一個脈沖在調節開始后的3-4秒施加,第二個脈沖在調節開始后的11秒之內施加。每次調節條件、試驗結束后,被試均獲得反饋(見圖2)。每次實驗均連續記錄瞳孔大小和心電圖。
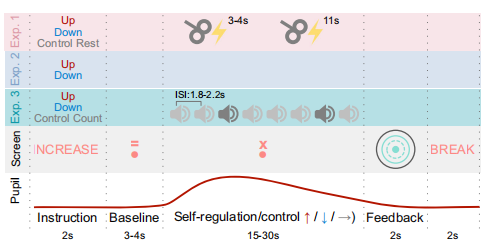
圖2 實驗1-3的流程
TMS參數設置
本研究使用單脈沖TMS,強度為120% RMT。在上調、下調或靜息控制條件下,分別在瞳孔調節階段輸出兩個TMS脈沖。第一個脈沖在調節(或休息)開始后3 - 4秒之間,第二個脈沖在11秒之后,以避免TMS脈沖預期。每個試次發送兩個脈沖,每種情況下收集40個MEPs樣本。
02 實驗二
實驗2將基于瞳孔的生物反饋與腦電圖相結合,研究瞳孔自我調節如何影響譜斜率(一種覺醒的皮層標記物)。本研究假設,與瞳孔縮小時(藍色)相比,在瞳孔放大(紅色)(表明更高的喚醒)期間,光譜斜率(皮質喚醒的全局標記)更平坦。右邊的條形圖顯示了估計的譜指數(圖3)。

圖3 以瞳孔為基礎的生物反饋與腦電圖相結合,研究瞳孔自我調節如何影響譜斜率
基于瞳孔生物反饋的瞳孔測量記錄
在這里,只報告基于瞳孔的生物反饋訓練第3天的數據,同時記錄了腦電圖和心電圖數據。在每次訓練中,受試者共進行60個試次,分為3個上調和3個下調block,每個block由10個試次組成。實驗開始時,對瞳孔大小的調節方向進行2秒的提示(放大或縮小瞳孔大小),然后進行3秒的基線測量。在接下來的實驗中,受試者在調節階段運用他們的心理策略來調節瞳孔直徑的大小。
腦電記錄
腦電圖記錄(均為德國Brain Products公司生產,深圳瀚翔腦科學技術股份有限公司全國總代理)。使用64導鹽水電極、actiCHamp Plus放大器和腦電信號記錄軟件BrainVision Recorder以1kHz的頻率記錄基于瞳孔的生物反饋過程中的腦電信號。使用actiCAP SNAP支架根據標準化的國際10-20系統將電極放置在腦電帽中。
03 實驗三
實驗3將瞳孔生物反饋與腦電相結合,研究瞳孔自我調節如何影響目標音調的譜斜率和P300事件相關電位(ERP)成分,這是藍斑(LC)活動的電生理標志物。研究假設瞳孔縮小(藍色)時的斜度比瞳孔放大(紅色)時的斜度更陡,并進一步研究斜度是否會隨著目標色調的變化而變化。此外,研究假設當受試者在目標音調之前瞳孔縮小(藍色)時,P300會比瞳孔放大(紅色)時更大。圖4中的聲音圖標代表在oddball任務中播放的標準(淺灰色)和目標音調(深灰色)(圖4)。
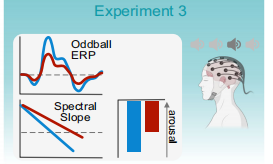
圖4 在oddball任務中播放的標準(淺灰色)和目標音調(深灰色)
研究結果
01 實驗一結果
盡管在調節階段施加了兩個經顱磁刺激脈沖,但研究受試者能夠自主地放大和縮小瞳孔大小(圖5)。瞳孔大小(圖6)在上調期間比下調時顯著增大。當受試者將自我調節策略應用于放大瞳孔時,皮質興奮性顯著增加了。
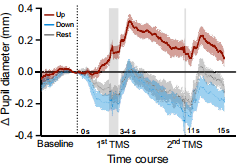
圖5 基于瞳孔的自我調節對皮質興奮性和心血管激活的影響
注:TMS 脈沖作用的時間進程,包括上調、下調和控制休息試驗。灰色陰影區域表示 TMS 脈沖的時間點,其中第一個脈沖在 3 - 4 秒之間施加,第二個脈沖在調節開始后 11 秒施加。每個試次都以基線測量開始(直到虛線),隨后是 15 秒的調節/休息階段,在此期間施加了 2 個 TMS 脈沖。
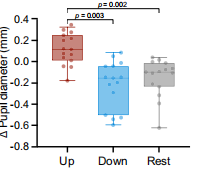
圖6 在每次經顱磁刺激脈沖前的 150 毫秒內,在上調、下調和對照條件下,瞳孔大小均有所變化。
注:在上調期間,修正后的基線瞳孔大小變化明顯大于下調和對照試驗組;下調期間的瞳孔大小變化沒有顯著低于對照組。
02 實驗二結果
正如假設的那樣,在瞳孔縮小期間的譜斜率明顯比瞳孔放大期間更陡峭(圖 7)。
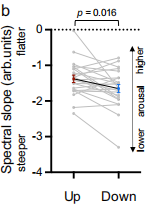
圖7 針對 30 - 45 Hz頻段所估算的光譜斜率,表明在瞳孔縮小時的斜率明顯比放大時的斜率更陡峭
對于 2 - 40 Hz范圍內估計的譜斜率的類似趨勢水平結果表明,在瞳孔縮小調節期間的皮質覺醒狀態低于瞳孔放大調節期間,且這種差異僅在自我調節期間存在,尤其是在自我調節開始后的 3 - 6 秒,而在基線期間不存在(圖8)。
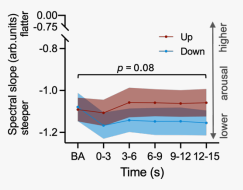
圖8 在基線(即-3-0s)和調節期間(0- 3s、3- 6s、6- 9s、9- 12s、12- 15s)的3 s時間區間內,所有上調和下調試次通道在30- 45hz的平均估計頻譜斜率。
瞳孔自我調節對腦電圖測量的全部皮質覺醒具有調節作用(圖9)。

圖9 在基線(即-3-0秒)和調節階段(0-3秒、3-6秒、6-9秒、9-12秒、12-15秒)的時間區間內,所有上調和下調試次在30-45 Hz的平均頻譜斜率估計。
瞳孔調節指數(上調 - 下調)與頻譜斜率變化(上調 - 下調)之間存在顯著的正相關,因此,更好的瞳孔自我調節與更大的頻譜斜率差異相關(圖10)。
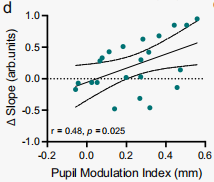
圖10 瞳孔和皮質覺醒標志物之間存在顯著的線性關系
03 實驗三結果
相比于上調和控制組,下調期間譜斜率更加陡峭(圖11)。
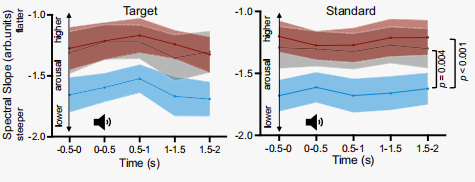
圖11 在30-45 Hz時所有通道向上、下調節和控制試次的光譜斜率
在下調和控制條件下,P300 的表現存在顯著差異(圖12-14),這種差異表現為,在整個中央頂葉區域,下調的幅度高于對照條件的調節幅度,并且這種差異在聲音開始后的 252 至 560 毫秒之間最為顯著,這與 P300 的預期時間特性和空間特性相吻合。同時,下調和上調條件之間的 P300 也有顯著差異。但是,P300在上調和控制條件之間沒有顯著差異。
通過比較P300對標準聲音和目標聲音的反應,進一步證實了瞳孔自我調節作用對目標誘發P300的特異性。總的來說,目標聲音在252 ~ 600 ms期間的平均振幅顯著高于標準聲音(圖14)。
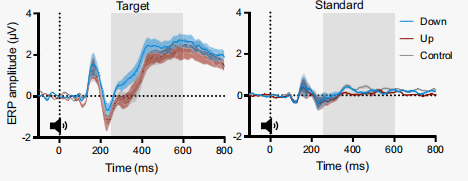
圖12 oddball任務期間的平均ERP波形
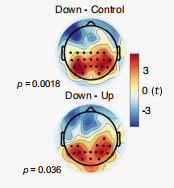
圖13 下調和控制組的P300存在顯著的差異
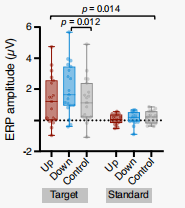
圖14 下調期間的P300振幅顯著高于控制組
研究結論
本研究將基于瞳孔的生物反饋與(i)單脈沖經顱磁刺激(TMS)、(ii)腦電圖(EEG)記錄以及(iii)同時進行的聽覺oddball任務相結合,揭示了三個主要發現:首先,瞳孔自我調節顯著影響了用 TMS 測量的皮質興奮性。其次,無論是在僅自我調節的條件下還是在自我調節與聽覺oddball任務同時進行的雙任務設置中,瞳孔自我調節均顯著影響了腦電圖功率譜的斜率(即頻譜斜率),這是人類皮質覺醒的電生理標志物,可通過捕捉興奮/抑制比的變化來反映內在興奮性。第三,瞳孔自我調節顯著影響了目標音調引發的 P300 反應。
總之,本研究結果表明,通過基于瞳孔的生物反饋訓練獲得的瞳孔自我調節能夠調節大腦皮層興奮性和喚醒度的標志物。尤其是瞳孔和皮質喚醒度測量之間的相關性支持了存在共同潛在生理機制的解釋。同樣,通過瞳孔自我調節對 P300 的系統性反應調節,補充了前人研究的瞳孔擴張隨瞳孔相關喚醒狀態變化的情況。未來的研究需要確定基于瞳孔的生物反饋是否能進一步用于調節與神經和精神疾病相關的方面。
聲明
本文由科教支持部整理,歡迎批評指正。
原文鏈接:https://www.nature.com/articles/s41467-025-59837-5

 打電話
打電話

 微信
微信

 QQ
QQ

 產品中心
產品中心



